ESPCI web site
ESPCI web site



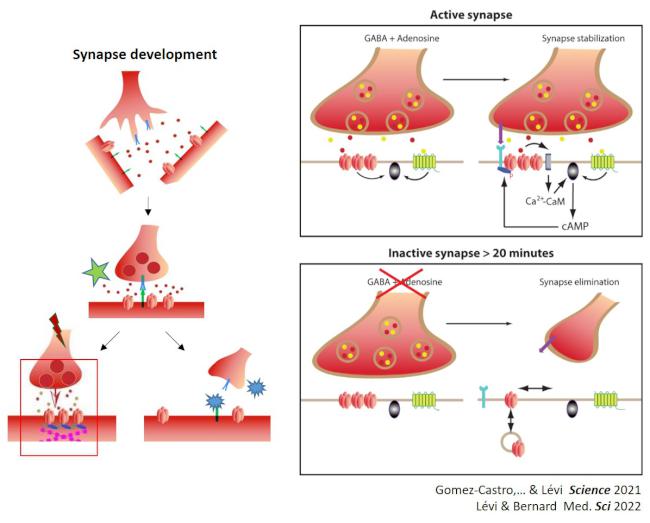
Développement : Nous avons montré que les récepteurs A2A de l’adénosine contrôlent le destin des synapses GABAergiques naissantes : le récepteur de l’adénosine A2A agit comme un senseur d’activité pour stabiliser les synapses GABAergiques actives capables de libérer le GABA et l’ATP alors que les synapses inactives sont rapidement éliminées. Puis nous avons mis en lumière le mécanisme sous-jascent.
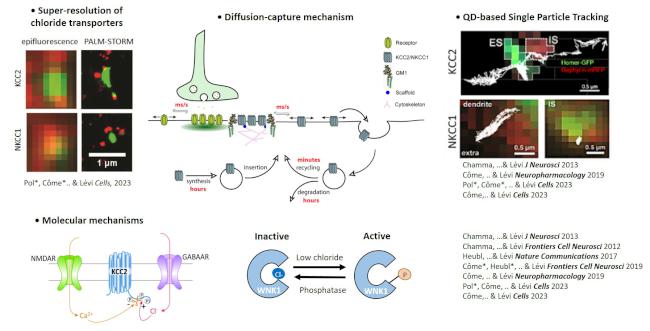
Plasticité des synapses matures : Ces dernières années, nous avons démontré l’importance du mécanisme de "diffusion-capture" dans la régulation rapide et plastique des récepteurs GABAA aux synapses inhibitrices de l’hippocampe ainsi que des transporteurs du chlore KCC2 et NKCC1 qui contrôlent l’homéostasie du chlore et ainsi l’efficacité de la transmission GABAergique. Puis, nous avons cherché des mécanismes de régulation de la diffusion-capture et ainsi de l’activité des récepteurs et des transporteurs du chlore. Cela nous a permis de mettre en évidence :
• L’impact des changements de conformation du récepteurs GABAA en conditions physiologiques (e.g. lors de l’ouverture, désensibilisation ou fermeture du canal) ou pathologiques (mutation humaine associée à l’épilepsie) ou de la géphyrine, la principale protéine d’échafaudage à la synapse inhibitrice (e.g. par phosphorégulation) ;
• Des comportements dynamiques différents entre KCC2 et NKCC1 et les mécanismes moléculaires permettant une régulation rapide de leur stabilité membranaire et ainsi de l’homéostasie du chlore neuronal ;
• Dans la recherche de nouveaux mécanismes de contrôle de la synapse inhibitrice, nous avons montré la contribution du chlorure comme messager secondaire capable d’activer les kinases WNK/SPAK, jusqu’ici peu explorées dans le cerveau, et qui agiraient en chef d’orchestre aux synapses inhibitrices en régulant de manière concomitante KCC2 et NKCC1. Cela fait de cette voie une cible d’intérêt dans les pathologies avec des altérations de l’inhibition comme l’épilepsie, les maladies psychiatriques et le stress.